Có nhiều yếu tố nguyên nhân khác nhau của động kinh như thiếu sót của sự tưới máu, những biến đổi hóa sinh do thiếu oxy, di chứng viêm não, u não, dị dạng mạch máu não, chấn thương sọ não,v.v….
Trên lâm sàng được gọi là “động kinh triệu chứng” hay “động kinh thứ phát”, biểu hiện lâm sàng rất dễ nhận biết, cơn cục bộ có thể trở nên toàn thể hóa, nhưng cũng có trường hợp từ một ổ đông kinh, do phóng lực quá mạnh và nhanh lôi kéo toàn bộ não bộ tham gia gây nên cơn toàn thể làm nhầm với cơn toàn thể thực thụ (hay cơn toàn thể nguyên phát).
Một số cơn co giật gặp ở trẻ nhỏ nhưng không thực là động kinh vì não của chúng chưa chín muồi, rất dễ nhạy cảm với những rối loạn chuyển hóa như giảm can xi huyết, giảm natri huyết, hạ đường huyết, sốt cao co giật,v.v…
CƠ CHẾ BỆNH SINH
Cơ chế bệnh sinh của động kinh rất phức tạp, để hiểu, nên điểm qua về nguyên lý hoạt động của nơron. Mỗi nơron gồm hai phần, thân có những đuôi gai và trục nơron bao quanh bởi một màng, màng đuôi gai có diện tích lớn hàng nghìn lần của thân và trục.
Sinh lý màng nơron
Mỗi nơron có hai cực gồm có cực tiếp thu các tín hiệu gồm thân và đuôi gai, một cực ra là sợi trục hay cực phát. Màng nơron có đặc tính ưu việt là thẩm thấu chọn lọc các ion trong và ngoài màng. Ion Na+ và Cl- tập trung ngoài màng, và ngược lại là ion K+ ở trong màng.
Tín hiệu thần kinh
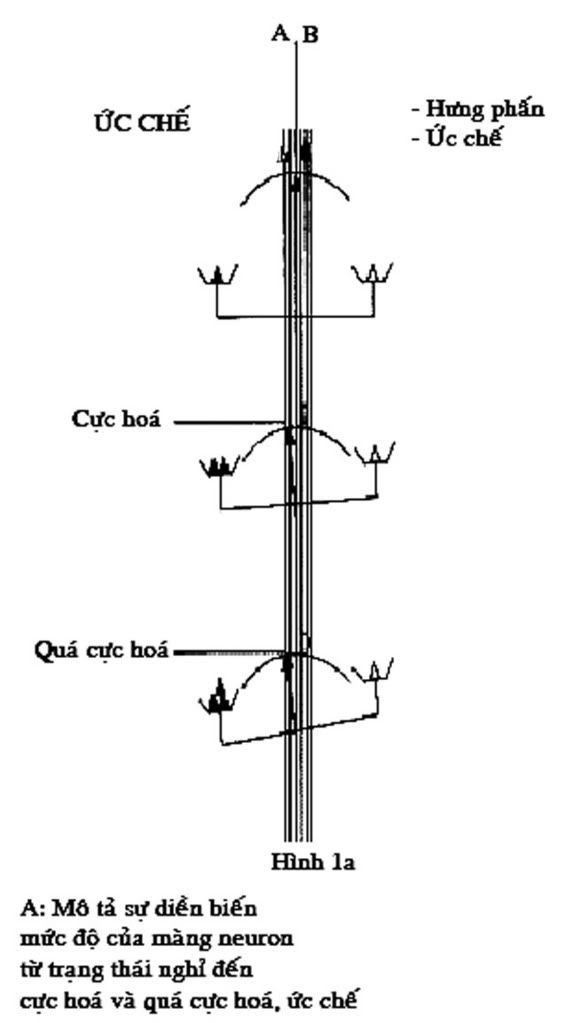
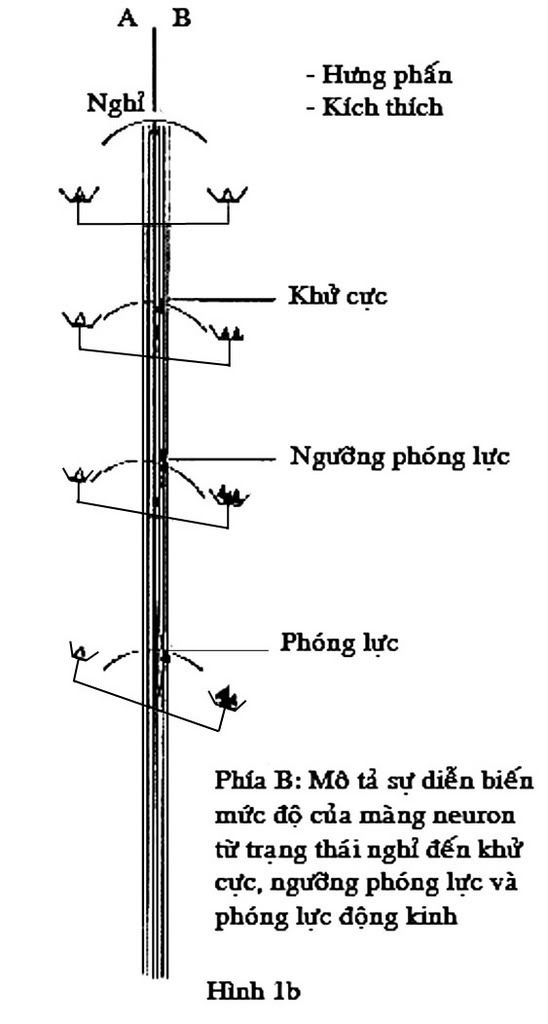
Một nơron bao giờ cũng có một điện thế màng, lúc nghỉ ngơi không hoạt động, bên ngoài màng luôn luôn dương so với bên trong âm, tạo nên điện thế là 70mV, như vậy nơron đang ở trạng thái cực hóa. Một tín hiệu bất kỳ nào đó có khả năng làm giảm điện thế xuống 10mV lập tức ở đó tạo nên một kênh để ion Na+ đi vào trong và ion K+ đi ra ngoài, màng lúc này bị khử cực và xuất hiện một dòng điện tại chỗ.
Khi sự khử cực của màng đuôi gai và thân nơron tới chỗ tiếp nối với sợi trục sẽ có hai khả năng xảy ra : sự khử cực chỉ đến đó rồi ngưng, tín hiệu coi như không truyền nổi; ngược lại, nếu sự khử cực vượt qua được để đi vào chỗ bắt đầu của sợi trục để rồi tới đoạn cuối của sợi trục sự truyền tín hiệu đã được thực hiện.
Sự dẫn truyền các tín hiệu thực hiện tại sinap thần kinh
Sinap gồm màng của túi tận cùng của sợi trục, còn gọi là màng tiền sinap, trong chứa nhiều bọng, trong đó là những chất dẫn truyền thần kinh khác nhau. Khe sinap là một khe khoảng trống nhỏ, tiếp đến là màng của đuôi gai và thân nơron thứ hai còn gọi là màng hậu sinap.
Khi sự khử cực xảy ra tại màng tiền sinap giải phóng các chất dẫn truyền thần kinh vào khe sinap chúng đi về phía màng hậu sinap, kích thích màng và màng bị khử cực, như vậy tín hiệu đã được thực hiện trót lọt từ nơron thứ nhất sang nơron thứ hai.
Hưng phấn và ức chế
Khi màng của nơron tiếp thu đang ở trạng thái khử cực nhẹ thì chỉ một tín hiệu yếu từ một nơron đang hoạt động truyền đến cũng đủ gây nên sự khử cực của màng nơron tiếp thu, như vậy màng ở trạng thái hưng phấn.
Ngược lại một tín hiệu đi đến màng hậu sinap, trong khi màng này ở trạng thái quá cực hóa, nó không gây nổi sự biến đổi của màng, có nghĩa nơron đang ở trạng thái ức chế.
Cơ chế gây động kinh
Động kinh là hậu quả của một sự phóng lực mạnh và đồng thời của một nhóm lớn những nơron bệnh lý. Chiều hướng nghiên cứu sinh bệnh lý động kinh hiện thời tập trung vào cơ chế của sự cân bằng và mất cân bằng của quá trình kích thích và ức chế của nơron (hình 1a + 1b).
Người ta có thể phân biệt chủ yếu hai loại dẫn truyền thần kinh : glutamate và acide gamma – amino butyrique (GABA). Glutamate là chấtdẫn truyền thần kinh (DTTK) kích thích, khi gắn lên những thụ thể glutamate lực thì gây nên một sự khử cực của tế bào đích. Ngược lại sự gắn của GABA, một chất DTTK ức chế, trên những thụ thể GABA lực gây nên một sự quá cực hóa của màng.
GABA là một chất DTTK ức chế chủ yếu của hệ thần kinh trung ương, khi nó được giải phóng vào khe sinap sẽ đi về hướng màng hậu sinap và gắn trên hai thụ thể GABA gồm thụ thể GABA loại B và thụ thể GABA loại A.
Thụ thể GABA loại B thuộc họ các thụ thể chuyển hóa dinh dưỡng (metabotrope), có cấu trúc gần giống với cấu trúc của thụ thể chuyển hóa glutamate. Thụ thể GABA loại B khu trú tại tiền sinap và hậu sinap, và có tác dụng gây nên một đáp ứng ức chế chậm bằng cách tăng độ dẫn điện của những kênh ion K+.
Thụ thể GABA loại A liên quan nhiều đến cơ chế cơ bản của động kinh, là đích nghiên cứu quan trọng của nhiều loại chống động kinh , và thuộc họ những thụ thể ion dinh dưỡnG (iontropes) vốn có một phần vai trò trong việc sát nhập các protêin.
HAI THỤ THỂ KÍCH THÍCH - THỤ THỂ AMPA VÀ THỤ THỂ NMDA
Glutamate là chất dẫn truyền thần kinh có khắp nơi trong hệ thống thần kinh trung ương, và có đặc điểm là tự gắn trên nhiều loại thụ thể của các kênh khác nhau. Có hai thụ thể chính là acide alpha-amino-3-hydro-5-methyl-4-isoxazoleproionique (AMPA) và N-méthyl-D-aspartate (NMDA). AMPA và NMDA là những phân tử đồng vận đặc biệt của hai thụ thể này.
Khi glutamate gắn vào thụ thể AMPA kênh sẽ mở ion Na+ đi vào bên trong và gây nên sự khử cực màng nơron gây nên một dòng điện rất ngắn. Cách hoạt động của thụ thể NMDA phức tạp hơn, khi glutamate gắn trên thụ thể này kênh chỉ mở nếu màng của nơron đã khử cực một cách đầy đủ. Cụ thể là khi màng có điện thế nghỉ làm cản trở sự lưu thông của các ion.
Nếu glutamate gắn lên thụ thể và gây nên được một sự khử cực đầy đủ thì ion Mg+, Ca++ đi vào và tạo nên một dòng điện có thời gian dài hơn so với dòng của sự hoạt hóa các thụ thể AMPA nói trên. Nguyên tắc chung là sự dẫn truyền glutamate lực có nhiệm vụ chủ yếu là hoạt hóa nhưng thụ thể AMPA trong tổ chức của hãi mã (hippocampe).
Những thụ thể NMDA chỉ can thiệp trong những điều kiện rất đặc biệt. Trong hiện tượng uyển chuyển (plasticité) của sinap, nó đóng vai trò quan trọng trong trí nhớ, và học tập. Khi những kích thích lập đi lập lại được thực hiện, màng nơron bị khử một cách đầy đủ để phá đi sự phong tỏa của ion Mg+. Khi Ca++ vào được trong nơron thì ion này sẽ gây nên một loạt những biến cố dẫn đến biến đổi sự đáp ứng của những thụ thể AMPA và NMDA.
Nguyên tắc chung là sự uyển chuyển của sinap là những thụ thể AMPA phụ thuộc vào sự hoạt động của những thụ thể NMDA, và thụ thể NMDA thì đóng vai trò trội trong các nhu mô động kinh, dĩ nhiên kể cả vai trò quan trọng trong bệnh lý động kinh và thiếu máu não cục bộ.
Sự không có oxy (anoxie) trong một khoảng thời gian gây nên một tiềm lực quan trọng của các thụ thể NMDA. Trong động kinh thái dương thụ thể NMDA tham gia một cách trực tiếp vào việc dẫn truyền sinap và việc kiểm soát sự phóng lực động kinh (hình 2).
NHỮNG VỊ TRÍ LIÊN QUAN ĐẾN VIỆC LÀM THAY ĐỔI SỰ CÂN BẰNG GIỮA ỨC CHẾ VÀ KÍCH THÍCH.
Sự mất cân bằng giữa hưng phấn và kích thích của mô động kinh có thể xảy ra ở nhiều nơi. Một mạch điển hình của các nơron thuộc diện CA1 hoặc CA3 của hải mã bao gồm một tế bào chính là tế bào tháp mà sợi trục của nó thì đi ra từ một vùng nhất định. Chất dẫn truyền của tế bào tháp này là glutamate vốn lại kích thích tiếp theo đó là hai nơron trung gian có chất dẫn truyền thần kinh là GABA ức chế.
Loại nơron trung gian ức chế 1 có sợi trục ù chỉ đi đến tế bào tháp, và thân của nó nhận những tín hiệu kích thích của tế bào tháp và những nơi khác. Loại nơron ức chế 2 có sợi trục chỉ đi đến nơron trung gian ức chế 1, không trực tiếp với tế bào tháp, do đó mà tín hiệu ức chế của nơron ức chế 2 chỉ được thực hiện qua trung gian của nơron ức chế 1.
Do đặc điểm trên mà nơron ức chế 2 được gọi là “nơron trung gian chuyên biệt của những nơron trung gian”. Từ sơ đồ này có thể xác định những điểm gây cơn với ý định giải thích sư sinh động kinh (hình 3).
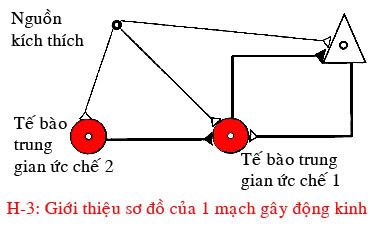
NHỮNG SINAP KÍCH THÍCH TRÊN NHỮNG TẾ BÀO CHÍNH
Những sinap này tăng sự hoạt động kích thích trên những tế bào tháp chính làm cho các nơron này đến gần ngưỡng sinh ra động kinh, sự gia tăng này gồm có :
Số lượng sinap kích thích gia tăng. Nhiều công trình thử nghiệm thấy rõ những sinap mới (tăng tiềm lực kích thích) hình thành nhiều trong mô động kinh ở người cũng như vật thí nghiệm. Những sợi rêu là những trục của những tế bào hạt của hồi răng và trong Stratum Oriens của điện CA 3.
Tăng tiềm lực lâu dài của những sinap kích thích. Sự tăng tiềm lực lâu dài là một hiện tượng được đặc tính hóa bởi việc gia tăng chắc chắn của sự đáp ứng của nơron hậu sinap. Nhưng sinap kích thích có thể tái tăng tiềm lực ngay sau khi nơron vừa mới phóng lực động kinh.
Thay đổi tiền và hậu sinap. Một thay đổi tiền sinap đưa đến sự phóng các bọng chứa glutamate vào khe sinap, hoặc số lượng nhiều hay ít số lượng những bọng được phóng sẽ gây nên những hậu quả quan trọng cho những đáp ứng hậu sinap. Một sự thay đổi sự chuyển dịch những thụ thể ngoài sinap về hướng hậu sinap cũng sẽ gây nên hệ quả quan trọng đối với sự cân bằng giữa kích thích và ức chế. Những luận chứng này cũng áp dụng được để giải thích sự hoạt động của những synapse ức chế đã nói ở trên.
NHỮNG THỤ THỂ NMDA VÀ ĐỘNG KINH
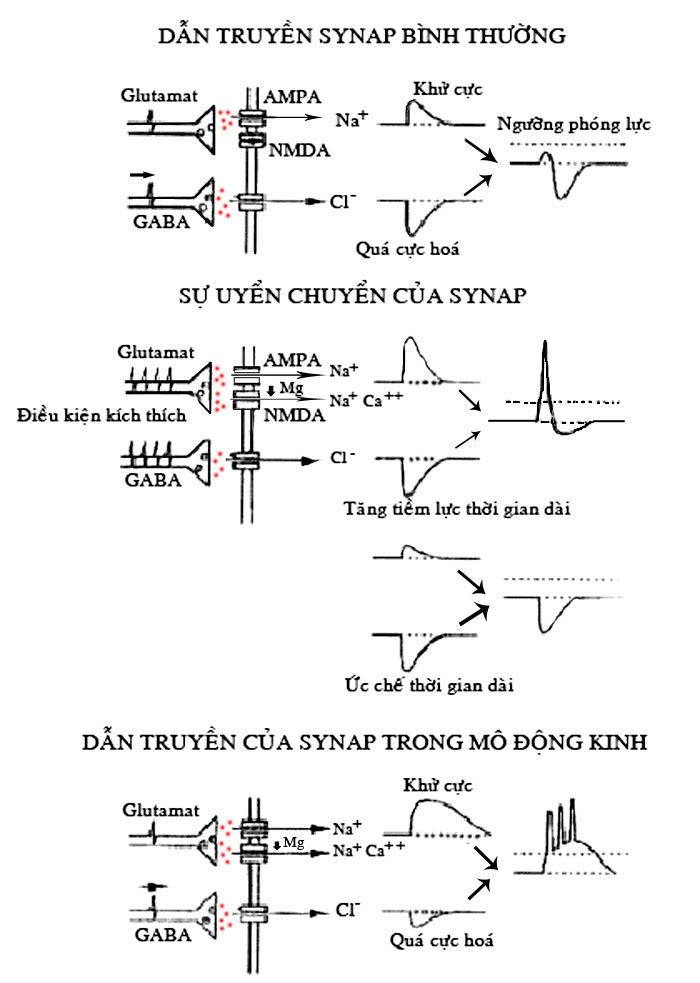
Sơ đồ dẫn truyền của Glutamat và GABA trong những điều kiện thực nghiệm khác nhau
Những thụ thể NMDA tham gia trực tiếp vào sự dẫn truyền sinap trong mô động kinh ở người và ở vật thí nghiệm, sự hoạt động của nó phức tạp hơn nhiều so với thụ thể AMPA vì nó có mặt tại nhiều vị trí có chức năng điều hòa hoạt động trong và ngoài tế bào, như vị trí glycine, polyamine, Zn++, pH và oxy hóa khử (redox) có khả năng kiểm soát hiệu quả và chính xác sự hoạt hóa của thụ thể NMDA.
Nhiều nghiên cứu cho thấy tác nhân oxy hóa khử 2-nitrobenzoic acid (DTNB) làm giảm 50% sự đáp ứng của thụ thể NMDA trong những nơron nuôi cấy. DTNB không có tác dụng trên các thụ thể AMPA và GABA lực. Đây là một hướng nghiên cứu đưa các thuốc này vào điều trị các bệnh lý thiếu oxy.
KẾT LUẬN
Động kinh đã trải qua hơn hai thiên niên kỷ, từ quan niệm thần bí nay đã được khoa học y học, sinh học phân tử, tìm hiểu một cách tương đối cặn kẽ, nhờ hiểu biết chức năng sinh lý của màng nơron, tiếp đến sinh lý, chức năng của sinap và các hệ thống lực của các chất dẫn truyền, người ta đi đến nghiên cứu cơ chế và bệnh sinh động kinh và đã rút ra những lý luận cơ bản :
- Động kinh là hậu quả của sự mất cân đối giữa hai quá trình ức chế và kích thích của nơron.
- Hoạt động ức chế là hậu quả của sự hoạt động của hệ thống GABA lực, chất dẫn truyền GABA, và các thụ thể GABA của màng hậu sinap.
- Hoạt động kích thích bắt nguồn từ những hệ thống glutamate lực, chất dẫn truyền là glutamate, thụ thể tương ứng có hai loại AMPA và NMDA các thụ thể này xuất hiện nhiều trong các tổ chức động kinh.
- Từ những hiểu biết về cơ chế sinh lý bệnh động kinh, người ta đã tìm ra những thuốc có tác dụng chọn lọc trên từng vị trí, trên từng thụ thể ví dụ một số thuốc chống động kinh chỉ có tác dụng với ổ động kinh cục bộ, có tác dụng ngược lại khi dùng cho các động kinh toàn thể không co giật. Cơ chế động kinh còn nhiều điều phải tìm hiểu hơn nữa ngõ hầu mới giúp ta quản lý, xử trí tốt căn bệnh này.
Những thụ thể NMDA tham gia trực tiếp vào sự dẫn truyền sinap trong mô động kinh ở người và ở vật thí nghiệm, sự hoạt động của nó phức tạp hơn nhiều so với thụ thể AMPA vì nó có mặt tại nhiều vị trí có chức năng điều hòa hoạt động trong và ngoài tế bào, như vị trí glycine, polyamine, Zn++, pH và oxy hóa khử (redox) có khả năng kiểm soát hiệu quả và chính xác sự hoạt hóa của thụ thể NMDA.
Nhiều nghiên cứu cho thấy tác nhân oxy hóa khử 2-nitrobenzoic acid (DTNB) làm giảm 50% sự đáp ứng của thụ thể NMDA trong những nơron nuôi cấy. DTNB không có tác dụng trên các thụ thể AMPA và GABA lực. Đây là một hướng nghiên cứu đưa các thuốc này vào điều trị các bệnh lý thiếu oxy.
KẾT LUẬN
Động kinh đã trải qua hơn hai thiên niên kỷ, từ quan niệm thần bí nay đã được khoa học y học, sinh học phân tử, tìm hiểu một cách tương đối cặn kẽ, nhờ hiểu biết chức năng sinh lý của màng nơron, tiếp đến sinh lý, chức năng của sinap và các hệ thống lực của các chất dẫn truyền, người ta đi đến nghiên cứu cơ chế và bệnh sinh động kinh và đã rút ra những lý luận cơ bản :
- Động kinh là hậu quả của sự mất cân đối giữa hai quá trình ức chế và kích thích của nơron.
- Hoạt động ức chế là hậu quả của sự hoạt động của hệ thống GABA lực, chất dẫn truyền GABA, và các thụ thể GABA của màng hậu sinap.
- Hoạt động kích thích bắt nguồn từ những hệ thống glutamate lực, chất dẫn truyền là glutamate, thụ thể tương ứng có hai loại AMPA và NMDA các thụ thể này xuất hiện nhiều trong các tổ chức động kinh.
- Từ những hiểu biết về cơ chế sinh lý bệnh động kinh, người ta đã tìm ra những thuốc có tác dụng chọn lọc trên từng vị trí, trên từng thụ thể ví dụ một số thuốc chống động kinh chỉ có tác dụng với ổ động kinh cục bộ, có tác dụng ngược lại khi dùng cho các động kinh toàn thể không co giật. Cơ chế động kinh còn nhiều điều phải tìm hiểu hơn nữa ngõ hầu mới giúp ta quản lý, xử trí tốt căn bệnh này.
Nguồn: thankinhhoc.com




Đăng nhận xét